![]()
| This is the reversetranscriptase domain . It is the only domain found in all retrotransposable elements. | |||
![]()
| This is the endonuclease domain. All retrotransposons with this domain are site specific. The active site of this domain has homology to some restriction endonucleases. | |||
| This is the endonuclease domain. It has homology to apurinic endonucleases. Retrotransposons having this domain are usually not site specific but R1 is highly specific for its 28s target site | |||
| The R2/28s RNA is shown assembled into a ribosomal subunit with the R2 portion of the co-transcript positioned outside the protein/RNA complex | |||
| Here the R2 protein can be translated from the exported co-transcript. | |||
| Here the R2 protein has entered the nuleus and bound the 3' UTR of the R2 co-transcript. | |||
| The R2 protein now bound to the R2/28S co-transcript has nicked the target site on the chromosome and, using the bound RNA as a template, has begun to reverse transcribe the R2 sequence onto the chromosome using the exposed the 3' hydroxyl to prime the new cDNA strand. | |||
|
Research overview |
|||||||||
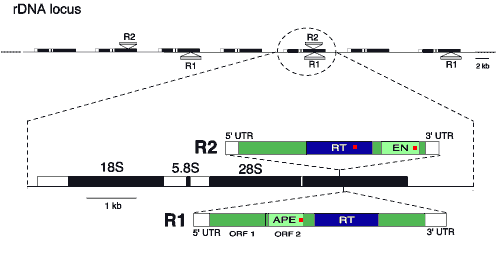
Transposable elements play a significant role in determining the size, structure and expression of eukaryotic genomes. For example, four million transposable elements constitute 45% of the human genome (175 elements for every gene). In all organisms a "genomic war" is being waged as mobile elements attempt to expand their copy number and the host organisms attempt to limit that number. The Eickbush laboratory is trying to understand this war on one localized battlefront. One of the most conserved and highly expressed gene families is the rRNA genes. All eukaryotes contain 100's to 1000's of tandemly arranged genes encoding the RNA components of the ribosomes. The organization of the 18S, 5.8S and 28S genes within this basic repeat (see Figure) and the function of these RNAs are identical across eukaryotes. The tandemly arranged rRNA genes, called here the rDNA loci, form the nucleoli observed in all cells. Little sequence variation is detected in the different copies of the rRNA genes present within a species. However, the sequences of all the genes change as a group over time, a process termed concerted evolution. The recombinational processes of concerted evolution, unequal crossovers and gene conversion, allow variants that are neutral or beneficial to become fixed in all units in the locus, while detrimental variants are eliminated. Given its uniformity in sequence and its critical role in metabolism, it is surprising that the rDNA loci have become the niche for a number of transposable elements. The 28S rRNA genes of many taxa including Arthropods, Nematodes, Platyhelminthes, Tunicates, Cnidaria and Vertebrates have been shown to contain such elements. We study the R1 and R2 elements of arthropods (see Figure). These elements are members of the class of retrotransposons variously called non-LTR retrotransposons, LINEs, or retroposons. Although they share a common location in the genome, R1 and R2 are only distantly related, encode distinct endonuclease domains, and thus represent independent specializations for insertion into the 28S gene. Because their insertion into 28S genes disrupts the expression of the entire unit, we initially assumed that selection forces and concerted evolution would frequently eliminate R1 and R2 elements from the rDNA loci of the host. However, R1 and R2 have been found in all lineages of arthropods examined. Furthermore, phylogenetic analyses have provided few examples of the loss of these elements from a lineage, and no evidence for transfers between species. We are left with the remarkable conclusion that these elements have been able to maintain themselves in all insect lineages since the origin of arthropods despite the fact they insert into a locus that is highly adapted to rid itself of variation. Our studies of R1 and R2 and the rDNA loci they inhabit are currently focused in three areas.
2. Population genetic studies of the concerted evolution of the rDNA loci. These studies involve the analysis of variation within populations, the utilization of the sequence data generated by genome sequencing projects, and the analysis of the changes that occur in laboratory strains over time. See section entitled Evolution of the rDNA locus. 3. Developmental and population genetic studies of the control over R1 and R2 expression. Strains of Drosophila have been identified with active and inactive elements. Control over that activity predominantly maps within the rDNA locus itself. Crosses between active and inactive lines suggest nucleolar dominance may play an important role in controlling element activity. See section entitled Regulation of activity. |
|||||||||
The rRNA genes of insects contain two retrotransposable elements inserted into the 28S gene. Top: view of a portion of an rDNA locus showing many of the tandemly repeated rRNA units with R1 and/or R2 insertions. Bottom: expanded view of one unit showing the 18S, 5.8S and 28S rRNA genes. The open reading frames of the R1 and R2 elements are shown containing similar reverse transcriptase (RT) but distinct endonuclease domains (APE and EN). |
|||||||||
|
Home |Mech of retrotransposition | Evolution of the rDNA locus |Regulation of acitivity| Publications | Members | |