Nasonia
![Nasonia Biology
Introduction
Nasonia are excellent organisms for research and teaching. These parasitoid wasps have been the subject of genetic, ecological, evolutionary and developmental research for over 50 years. Two general features that make these insects such excellent study organisms are (a) ease of handling and rearing, and (b) interesting and diverse biology. Nasonia are readily reared on commercially available fly pupae (the hosts). Virgin females and males are easily collected in the pupal stage (there is a 3 day time window for virgin collection). Adults are "user friendly" and can be handled without the need for anaesthetization. Nasonia has a short generation time (two weeks), but can be stored under refrigeration for periods of time, allowing for flexibility in experimental timing. A diapausing larval stage allows storage of strains for up to two years without maintenance. Both visible mutants and molecular markers are available for genetic mapping and instruction in genetics.
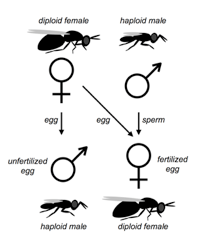
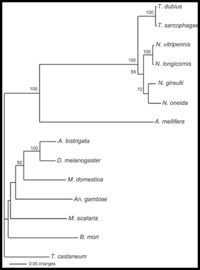
The system is excellent for basic studies in genetics, ecology, behavior, development and evolution. Four closely related species of Nasonia are present. The species are interfertile, allowing movement of chromosomal regions (and phenotypes) between the species for genetic and molecular genetic analyses of species differences in behavior, development, morphology and physiology. Nasonia is an excellent candidate for comparative genomic studies, as well. A key feature of Nasonia is haplodiploid sex determination; males are haploid and develop from unfertilized eggs and females are diploid and develop from fertilized eggs. This feature makes Nasonia a very useful organism for genetic research (advantages of this feature are described further below). Below we describe the basic biology of Nasonia, and discuss opportunities for research.
Basic Biology
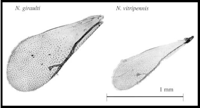
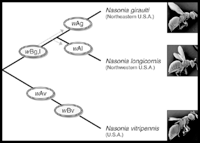
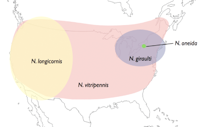
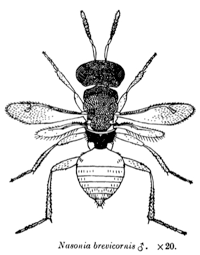
Nasonia are small parasitoid wasps (Hymenoptera: Pteromalidae) that sting and lay eggs in the pupae of various fly species, primarily blowflies and fleshflies. There are four closely related species in the genus, N. vitripennis, N. longicornis, N. giraulti and N. oneida. N. vitripennis is found throughout the world; N. giraulti is found in eastern North America; N. longicornis is found mainly in western North America and N. Oneida, a recently described species, is so far found only in upstate New York. Their approximate distribution in North America is shown in the accompanying distribution map. There are many intriguing aspects to Nasonia biology. Below we outline some of the basic features. A dated, but still excellent review of Nasonia biology is present in Whiting (1967).
Life Cycle
The basic life history described below is for N. vitripennis; the other three species have similar life-histories, and differences between the species will be mentioned.
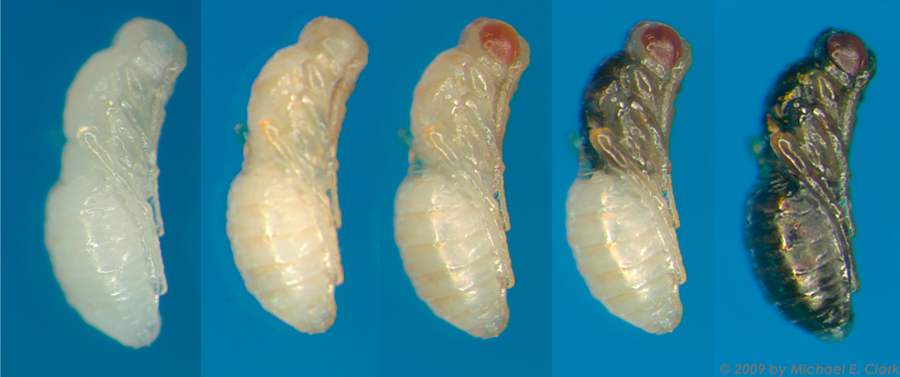
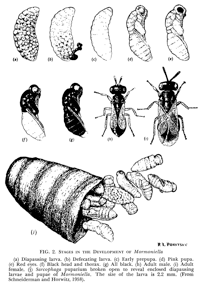
When a female encounters a host puparium, she first examines the host, then drills through the host puparial wall with her ovipositor. She injects venom into the pupa, which will eventually kill the fly. The female then commences laying eggs upon the host, underneath the puparial wall. She typically lays from 20 to 50 eggs per Sarcophaga bullata pupa. The female may lay these eggs in one bout or may take a number of hours to complete oviposition. The female also uses excretions from her ovipositor to construct a feeding tube from the pupa to the puparial wall. From this she feeds on host hemolymph, which appears to be important in the production of additional eggs. At 25°C, eggs hatch around 36 hours after being laid. Developing larvae complete 3 instars and then pupate within the host around 9 days after laying (see Development Time Table). Pupal development takes approximately 3 days. Male and female pupae can easily be distinguished during this time. Adults eclose from pupation within the host, and then chew an exit hole. Emergence typically occurs by 14 days.
Mating occurs immediately upon emergence from the host. Courtship behavior is brief (typically taking 1-2 minutes) and involves stereotypic courtship displays. After mating, females disperse from the natal patch in search of new hosts.
Development takes slightly longer in N. giraulti, N. longicornis and N. oneida. N. giraulti females often mate within the host prior to emergence, in contrast to the other species.
Genetics
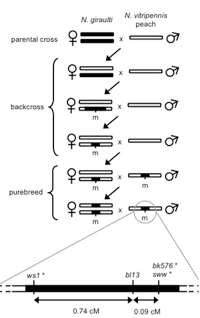
In many respects, Nasonia is a superior organism for genetic research. The important features that make it so are (a) short generation time (b) large family sizes, (c) ease of handling (including virgin collection), (d) ability to inbreed and produce healthy inbred isogenic lines, (e) availability of visible and molecular markers (f) ease of complete genome screening for mutations in the haploid sex, (g) presence of four closely related and interfertile species, which provides a wealth of phenotypic and molecular marker differences, and (h) ability to produce hundreds of genetically identical (clonal) recombinant genotypes in the F3 generation (see description below). These features make Nasonia an excellent organism for basic studies in genetics, including developmental genetics, evolutionary genetics, molecular evolution and comparative genomic research. Nasonia is particularly suited for the study of complex genetic traits, due to advantages provided by haploid males and the ability to easily produce inbred lines and genetically identical recombinant individuals. Positional cloning is practical in Nasonia, due to the high recombination rate and abundance of molecular marker differences between the interfertile species.
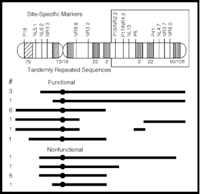
Basic Genetics: All Nasonia species normally have 5 chromosomes, corresponding to 5 linkage groups. A visible mutant map of Nasonia exists; currently there are about 20 mutant strains available, most of which are eye color, body color, morphological and embryonic lethal mutations (Saul 1989). Screening for new mutations in Nasonia is straightforward, given that the complete genome can be screened for recessive mutations in the haploid sex. The generation and characterization of new mutations is definitely needed in Nasonia. A more complete visible mutant map will be useful in genetic and developmental genetic studies, and will also facilitate positional cloning studies. In addition, screening and mapping of de novo mutations in Nasonia are practical projects for undergraduate researchers, who have the opportunity to discover new mutations in this system. There are also interesting opportunities for characterization of existing mutations in Nasonia. For example dant, (distal antennapedia) is a recessive homeiotic mutation that converts antennae to legs; it has not been extensively characterized, nor has it been determined whether this mutation is homologous to antennapedia in Drosophila.
In addition to a visible mutant map, a RAPD molecular map (Gadau et al 1999) and AFLP marker map (unpublished) have recently been generated. Production and mapping of molecular markers in Nasonia is surprisingly easy. This is because there is a high incidence of sequence differences between the species, and polymorphisms can be quickly mapped in haploid F2 males without the problems of dominance that can occur with many molecular markers. In addition, a set of hybrid recombinant inbred lines are coming available to use for even more rapid mapping of molecular markers. Quantitative Trait Locus studies are very feasible in Nasonia, particularly for traits in haploid males (see below)
Molecular Genetics & Comparative Genomics: The genome size of Nasonia vitripennis is approximately 250 Megabase (2X greater than Drosophila melanogaster); however, the recombination rate in Nasonia is approximately 4X greater than in D. melanogaster, resulting in an average recombination rate per kilobase approximately 2X greater (around 330 Kb/cm). This coupled with the ease of generating molecular markers suggests that positional cloning is practical in Nasonia. However, this has not yet been demonstrated.
There has been virtually no work done on topics such as gene regulation and expression in Nasonia, except for recent promising studies of early patterning mutants (described below under development). Some work has been conducted on repetitive DNA in Nasonia (Eickbush et al 1992) and a family of retrotransposable elements have been partially characterized in Nasonia (McAllister and Werren 1997).
Little is currently known about sequence differences within and between the Nasonia species. This is a research area with good potential. In addition, when a particular sequence difference has been identified, it can can be quickly mapped using recombinant F2 males or hybrid inbred lines. Therefore, Nasonia is a good candidate for comparative genomic studies in insects. This method has been used to map several genes involved in the insulin signaling pathway, indicating potential of the approach.
Developmental Genetics: Nasonia provides interesting contrasts to the standard insect model for developmental genetics, Drosophila melanogaster. These features are described below in the Development section.
Evolutionary & Quantitaive Genetics: Given the existence of closely related and interfertile species, opportunities for evolutionary genetic studies are abundant. Strains have been collected from different populations in North America for all three species, and these are available for laboratories interested in population genetic research. Analysis of mitochondrial CO1 sequences suggests some population subdivision in N. giraulti and N. longicornis. Studies are currently underway to characterize some phenotypic differences (e.g. wing size and female mate preference) between the species. The tools for detailed evolutionary genetic studies are now in place, and this promises to be a growth area in the near future.
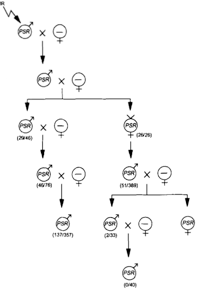
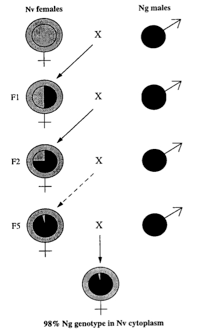
In Nasonia, epistatic gene interactions can more easily be investigated without the added complexity of dominance interactions, by using haploid males. The ability to produce isogenic inbred lines in Nasonia is a further advantage for quantitative genetic studies, since isogenic females can be placed in different environments to investigate genotype x environment interactions and norms of reaction. Finally, there is a feature fairly unique to Nasonia, which makes it very useful for quantitative and other genetic studies. Crosses can be performed between strains (or species) with different phenotypes; virgin F1 females are then provided with hosts. Because of haplodiploid sex determination, these females produce recombinant haploid male progeny. Individual males are haploid and therefore produce identical haploid sperm. Therefore, recombinant haploid males can be crossed to inbred line females, and the resulting F3 females will all be genetically identical (clonal females), but with a recombinant genotype. This permits, in the F3 generation of a cross, the production of hundreds of genetically identical females for analysis. Genetically identical recombinant females can be placed in different environments to analyze genotype x environment effects. In addition, F2 males can mate with many dozens of females, allowing crossing of the same haplotype into many different genetic backgrounds, each then producing hundreds of females for phenotypic characterization. The F2 recombinant males can readily be genotyped (e.g. using molecular markers) without marker codominance problems, and the genotype of the F3 females is known by also genotyping the maternal inbred line. These features make Nasonia almost uniquely adapted (among higher eukaryotes) for the study of complex genetic traits.
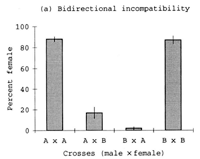
An exciting feature of Nasonia speciation is the presence of Wolbachia, cytoplasmically inherited bacteria that cause sperm-egg incompatibilities. All three species of Nasonia typically harbor two strains of Wolbachia, and these induced a high level of reproductive incompatibility between the species. This topic has been the subject of considerable research (e.g Breeuwer and Werren 1990, Bordenstein et al 2001). In fact, it is antibiotically cured strains of Nasonia that are used in interspecies crosses. These allow introgression of genes between the species, once the bacteria have been eliminated.
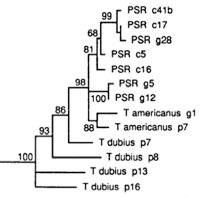
Although F1 females have high survival and fecundity, F2 hybrid males suffer from increased mortality during development. Taking advantage of the haploid genotypes of these males, a number of pairwise epistatic interactions have been mapped that lead to F2 male mortality (Gadau et al 1999). However, the developmental basis of F2 hybrid mortality has not been studied in detail.
Ecology & Behavior
Nasonia is an interesting organism for behavioral and ecological research. Its parasitoid life style allows investigations of questions relating to parasitoid-host dynamics, host preference, specialist versus generalist biology, et cetera. In terms of behavior, there are many interesting questions about courtship behavior, male aggression and territoriality, female dispersal, and sex ratio control. Presence of three closely related species with different biologies is useful, particularly because they are interfertile which allows movement of genes involved in these phenotypes between the species.
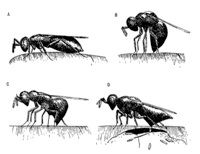
Courtship and Mating: Courtship involves stereotypic displays that differ between the species (van den Assem and Werren 1994) as well as the release of pheromones from the males mandibular region that plays an important role in female receptivity (van den Assem et al 1980). The Courtship display of N. longicornis is shown at the indicated link. Courtship occurs quickly (typically it is completed within 1-2 minutes) making it a tractable subject of study in undergraduate laboratories and for undergraduate research.
The genetic basis of courtship differences between the species is tractable for genetic analysis because of the ability to move genes between the species by hybridization and back-crossing of the fertile hybrids. Females of N. giraulti often mate within the host, whereas this is less common or absent in the other species. Within-host mating clearly will have strong influences upon the population structure. Males show territorial behavior, defending host puparia that have female wasps within. Little work has been done on this interesting behavior. After mating, females disperse from the natal patch in search of new hosts. Dispersal behavior of females differs between strains and species. Males of N. vitripennis have vestigial wings and are incapable of flying. Males of N. longicornis have intermediate sized wings and N. giraulti males have large wings similar in size to those of females. The latter two species are capable of flying, although they do not do so as readily as females.
Sex Ratios and Sex Ratio Distorters: Most matings occur locally within the natal patch, and sibling matings are not uncommon. Therefore, Nasonia is subject to local mate competition, and has been shown to alter sex ratio among progeny in response to the number of females in a group of hosts or as a consequence of superparasitism in patterns consistent with local mate competition theory (Werren 1980, 1983, but see Parker and Orzack 1985, Orzack and Parker 1986). When ovipositing, single females typically produce strongly female-biased sex ratios (80 -95% daughters), whereas when in groups they produce more equal ratios. Presumably, the haplodiploid sex determination provides a mechanism for control of the sex ratio among offspring, and reproductive anatomy of females suggest that they can control individual fertilization of eggs (Whiting 1967).
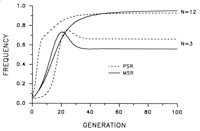
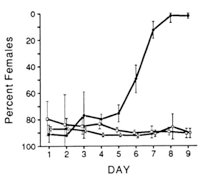
In addition to the normal sex ratio control of the wasps, a suite of extrachromosomal sex ratio distorting factors exist in natural populations. These include psr (paternal sex ratio), a supernumerary chromosome that causes destruction of the paternal chromosomes following fertilization, resulting in conversion of males to females, son-killer, a bacterium that kills unfertilized (male) eggs of infected females, msr (maternal sex ratio), a cytoplasmic factor that causes nearly 100% fertilization of eggs. These factors are maintained in different lines of Nasonia, allowing for detailed biological study.
Host Preferences: The different Nasonia species differ in their host preferences. N. vitripennis is a generalist and will parasitize a wide range of fly hosts, including blowflies, fleshflies and houseflies. The other three species appear to be specialists, and are found parasitizing Protocalliphora, blowflies that specialize as ectoparasites in bird nests. N. giraulti and N. longicornis prefer these hosts, although they will parasitize S. bullata. The behavioral, genetic basis of host preference differences has not been well studied.
Field Biology: Nasonia is a tractable, although occasionally smelly, system for field research. Wasps can be collected from bird nests and from the vicinity of carcasses (N. vitripennis). Baits using meat that has been fed upon by blowfly larvae placed in mesh bags can be efficiently used to sample natural populations. Field studies have uncovered a variety of the important features of this system, including sex ratio distorters, additional species, and strain differences in behavior and morphology. Strains collected from throughout North America are available to interested researchers, as is more detailed information on field sampling techniques.
Development
Nasonia is a good candidate for comparative studies of development. Mutations disrupting development can be rapidly screened for in haploid embryos, and maintained heterozygously in females. Genes affecting development can be quickly mapped using visible markers and the abundance of molecular marker differences present between the closely related species. These marker differences and a high recombination rate also make positional cloning a practical possibility within Nasonia. Recent work has uncovered several mutations affecting early pattern formation that appear to be homologous to homeiotic mutations in Drosophila (Pultz et al 2000), and also indicate that zygotic control of early development is more prevalent in Nasonia (Pultz et al 1999).
Additional work involves studies of morphological and developmental differences between the three closely related species. For example, males of the three species differ significantly in wing size and head shape. Genetic analysis of these features is tractable, including the eventual positional cloning of genes involved in these species differences. Preliminary work indicates a relatively simple genetic basis to wing size differences (Weston et al 1999). Excellent opportunities exist for detailed studies of head development using the natural variation present in the three species.
Handling and Rearing
Nasonia is easy to work with. Below are some of the relevant features that make them convenient laboratory organisms.
Stock Maintenance: Stocks are easily maintained in Nasonia in plastic or glass vials or test tubes. Emerging females are collected into a new vial by placing the vial over the original vial with emerged wasps. Females are negatively geotaxic and move into the new vial. Hosts are then placed into the new vial (usually approximately 1 - 2 wasps per host). Fourteen days later (at 25°C) the next generation emerges. It's as simple as that. No special feeding or handling is necessary. Stocks can be slowed down by placing them at cooler temperatures, or speeded up (up to about 28°C). Cultures can be placed under refrigeration for a couple of weeks if necessary. This is best done at the yellow pupal stage and adult stage, but can be done at other life stages as well. Adult females can also be kept alive for several weeks at 25°C with a small amount of honey, and females can live for over a month if provided with fresh hosts.
Collecting Virgins: Virgin collection is very easy in Nasonia. Wasp pupae can be sexed in the pupal stage, which provides a three day time window for virgin collection. They are immobile in the pupal stage, and therefore can be collected without the need for anesthetization. Individuals are most easily sexed in the dark pupal stage, but with minimal training can be readily distinguished as yellow pupae. One looks for the presence of an ovipositor in the distal end of the abdomen. In N. vitripennis, males can also be distinguished by small wing pads.
Handling Adults: Adults are very "user friendly" and can be sorted and used in experiments without anesthetization. Although females can fly, they do not do so readily. However, they are positively geotactic. Therefore, to set up females individually on hosts, one need only dump a few females onto a surface and then place test-tubes over the crawling individuals. They will then conveniently climb into the tube. Add a host (or two) and plug the tube with cotton and you are done. Large numbers of individuals can be efficiently handled in this way.
Collecting Eggs: The easiest way to collect eggs is to allow females to lay eggs for a prescribed period of time on a host to which their access is restricted to one end. This is accomplished by placing the host into a foam plug with a hole in one end, and placing this with the female into a test tube. After an oviposition period (the narrower the time, the more synchronized the eggs), hosts are removed, the puparial end is "popped off" with a probe, and eggs are collected with a fine brush. For maximum egg production, it is recommended that females be allowed to host feed for 2 - 3 days prior to placing them onto "plug hosts" for egg collection.
Diapause: Diapause larvae can be stored under refrigeration for up to two years, and then removed to room temperature, where they will complete development. Although two years is possible, for safety it is recommended that cultures be removed from diapause after around 1.5 years. Induction of diapause is accomplished by placing ovipositing females into short photoperiods (6L:18 D) and cool temperatures (e.g 15- 180 C). Better results are achieved by providing females with new hosts every several days under these conditions, and by allowing females to oviposit individually in test tubes. On occasion, a few generations are needed prior to diapause induction, and strains differ in diapause tendency.
Obtaining Hosts:
In working with Nasonia, you also need to have hosts. Fortunately, these are easy to obtain and also to rear, if you prefer. Nasonia vitripennis can be maintained on a number of different species, including Sarcophaga bullata (the fleshfly), various calliphorid flies including Calliphora vomitora, C. vicina, Phormia regina, and Phaenicia sericata, and on houseflies (Musca domestica). N. longicornis and N. giraulti can be cultured on blowflies and calliphorid flies (although their preferred hosts are Protocalliphora bird nest flies).
Hosts can be purchased from Ward's Natural Science, Carolina Biologicial and various other sources. Blowfly larvae (referred to as "spikes") are used as bait by fisherman, and can be purchased by bait stores in some areas. sufficient numbers reared in one round to maintain wasps for several months.
Sarchophaga bullata pupae can be placed under refrigeration (4°C) for several months and remain suitable for parasitization. Host quality is checked by cracking open the puparium at the head region of a few hosts (to be discarded). Hosts are suitable up to the brownish eye stage, although are preferable when in the white-eye to yellow-eye stage. Once bristles begin to form on the body or the body begins to darken, the hosts are unsuitable.
References
Saul, G., and B. Ii. 1989. Gene map of the parasitic wasp Nasonia vitripennis (=Mormoniella vitripennis), pp. 198-201. In : O'Brien, S.J. (ed) Genetic Maps: locus maps of complex genomes, pp 198-201. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
Eickbush, D., T. Eickbush and J.H. Werren. 1992. Molecular characterization of repetitive DNA sequences from a B chromosome. Chromosoma 101:575-583.
McAllister, B.F., and J.H.Werren. 1997. Phylogenetic analysis of a retrotransposon with implications for strong evolutionary constraints on reverse transcriptase. Molecular Biology and Evolution 14:69-80.
Gadau, J., R. Page and J.H. Werren. 1999. Mapping of hybrid incompatibility loci in Nasonia. Genetics 153:1731-1741.
Bordenstein, S.R., F.P. O'Hara and J.H. Werren. 2001. Wolbachia-induced bidirectional incompatibility precedes other hybrid incompatibilities in Nasonia. Nature 409:707-710.
Breeuwer, H. and J.H. Werren. 1990. Microorganisms associated with chromosome destruction and reproductive isolation between two insect species. Nature 346: 558-560.
Werren, J.H.. 1980. Sex ratio adaptations to local mate competition in a parasitic wasp. Science 208:1157-1160.
Werren, J.H.. 1983. Sex ratio evolution under local mate competition in a parasitic wasp. Evolution 37: 116-124.
Parker, E.d., and S.H. Orzack. 1985. Genetic-variation for the sex-ratio in Nasonia-vitripennis. Genetics 110:93-105.
Orzack, S.H., and E.D. Parker. 1986. Sex-ratio control in a parasitic wasp, Nasonia-vitripennis .1. Genetic-variation in facultative sex-ratio adjustment. Evolution 40:331-340.
Whiting, A.R. 1967. The biology of the parasitic wasp Mormoniella vitripennis [=Nasonia brevicornis] (Walker). The Quarterly Review of Biology 42:333-406.
Pultz, M.A., K.K. Zimmerman, N.M. Alto, M. Kaeberlein, S.K.Lange, J.N. Pitt, N.L. Reeves, and D.L. Zehrung. 2000. A genetic screen for zygotic embryonic lethal mutations affecting cuticular morphology in the wasp Nasonia vitripennis. Genetics 154:1213-1229.
Pultz, M.A., J.N. Pitt, and N.M. Alto. 1999. Extensive zygotic control of the anteroposterior axis in the wasp Nasonia vitripennis. Development 126:701-710.
Weston, R., I. Qureshi and J.H. Werren. 1999. Genetics of a morphological difference between two insect species. Journal of Evolutionary Biology 12:586-595.](WerrenLab-NasoniaBiology_files/shapeimage_1.png)
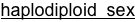

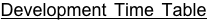
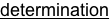
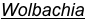
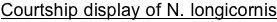
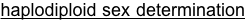









Nasonia


©2011 · University of Rochester , Department of Biology, Rochester, NY 14627 updated Jan 31, 2011